





CopyRight 2009-2020 © All Rights Reserved.版权所有: 中国海关未经授权禁止复制或建立镜像

双目峰口岸鼠类携带汉坦病毒情况分析
作者:王玮琳 徐阳春 杨迪 姜永莉 邹晓龙 洪晓坤 吴星 刘阳
王玮琳 徐阳春 杨迪 姜永莉 邹晓龙 洪晓坤 吴星 刘阳
Abstract This study is to grasp the distribution of rodents and the prevalence of Hantavirus at Shuangmufeng Port, so as to strengthen the prevention and control of Hantavirus in the port area. In the Shuangmufeng Port area, the mice were trapped by night-time method. The lung tissue of the samples was taken to extract RNA, RT-PCR detection of Hantavirus was carried out with specific hantaan primers (M segment), and the positive samples were isolated and purified, after the sequencing results were compared by NCBI-blast. MEGA7.0 was used to construct a partial M-segment Hantavirus phylogenetic tree (Neighbor-Joining method). A total of 97 mice of 5 species were collected, including Clethrionomys rutilus, Clethrionomys rufocanus, etc. All of which were Seoul Hantavirus (SEO), with a virus-carrying rate of 22.68%. The highest poisoning rate was 29.42%, followed by Clethrionomys rutilus and Clethrionomys rufocanus. The M-segment Hantavirus phylogenetic tree constructed based on the neighbor joining method (NJ) showed that all 22 SEO-type Hantavirus sequencing samples were clustered with Korean Hantavirus DPRK08. From this background monitoring, it was shown that the Clethrionomys rufocanus, the Clethrionomys rutilus, and the Apodemus peninsulae were the dominant species in the forest area, and the detection rate of the Hantavirus pathogen was relatively high.
Keywords Shuangmufeng Port; Hantavirus; evolutionary tree
汉坦病毒在世界范围内分布广泛,是重要的人畜共患病原体,给人类生产生活带来严重影响。汉坦病毒是布尼亚病毒科的成员,由三段负义单链RNA组成。第一个汉坦病毒分离株(TPMV),来自食虫动物的脾脏,于1964年在Vellore(印度)捕获[1]。亚洲汉坦病毒原型,汉坦病毒(HV)于1978年被分离出来,以捕获啮齿动物的汉江流域命名[2]。汉坦病毒经常引起肾综合征出血热(HFRS)和汉坦病毒心肺综合征(HCPS)两种急性发热疾病,中国由于内陆地域范围广阔,也是世界上受汉坦病毒危害较为严重的国家之一,全球90%以上的HFRS病例发生在中国[3]。
吉林省位于我国东北部,对外与朝鲜陆路接壤,对内与辽宁省、黑龙江省、内蒙古自治区相邻,是我国发现汉坦病毒较早且流行范围较广的省份之一[4]。我国吉林省安图县与朝鲜三池渊郡接壤,是发展环长白山旅游的最佳边境通道,而双目峰口岸是安图县境内中朝两国边界线上的陆路通道,因此,掌握双目峰口岸鼠类分布以及汉坦病毒流行情况对该地区HFRS的防治起着重要作用。
1 材料与方法
1.1 样本来源
选定双目峰口岸区域,用2号鼠夹,以花生米为诱饵,晚放晨收。采用夹夜法捕鼠,鼠夹间隔5 m,行间距50 m。2017年7月放置900个鼠夹,9月放置600个鼠夹,将捕获的鼠在生物安全二级实验室内解剖,取鼠肺置于液氮中保存。
1.2 方法
1.2.1 鼠脏器的研磨
将鼠肺标本从液氮中取出,取黄豆粒大小鼠肺置于2 mL 离心管内(管内2 个3 mm 氧化锆珠),加入500 μL 胎牛血清,用组织研磨器(Tissuelyser,Qiagen,德国)研磨(50 Hz,4 min),将组织研磨成匀浆。将匀浆液4℃,1000 r/min,离心5 min,上清转至1.5 mL 离心管中,待进行核酸提取。
1.2.2 鼠组织总RNA提取及cDNA制备
使用QIAamp Viral RNA Mini Kit(Qiagen,德国)提取鼠肺总RNA,按说明书操作。使用PrimeScript Ⅱ 1st Strand cDNA Synthesis Kit(TaKaRa)试剂盒将提取的RNA逆转录成cDNA。
1.2.3 汉坦病毒引物序列及PCR检测
汉坦病毒引物序列引用于《全国肾综合征出血热监测方案》(2005),见表1。
PCR反应体系:Premix Ex Taq™ Hot Start Version 12.5 μL、引物各0.5 μL(20 pmol/μL)、cDNA 3 μL,去离子水8.5 μL,总反应体系为25 μL。PCR反应条件:95℃预变性10 min;94℃ 1 min、55℃ 1 min、72℃ 45 s,共35个循环;72℃,延伸12 min。PCR产物通过1.5%琼脂糖凝胶电泳进行检测。阳性样品采用DNA胶回收试剂盒进行纯化,将纯化后的产物委托大连生工生物工程有限公司测序。
1.3 汉坦病毒系统发育树的构建
测序数据经Blast比对后,采用MEGA 7.0软件对测序数据构建系统发育树[5]。在NCBI数据库中下载相关参考汉坦病毒M段基因序列,采用邻接法(NJ)构建汉坦病毒部分M段进化树[6-8]。
2 结果与分析
2.1 双目峰口岸鼠类样品采集及鼠肺样本检测结果
双目峰口岸地区共捕获鼠97只,检测鼠肺样本97份,其中检出汉坦病毒阳性22例,带毒率22.68%。在汉坦病毒阳性的鼠中,大林姬鼠带毒率最高的为29.42%,其次为红背䶄和棕背䶄,带毒率分别为28.57%、16.33%;虽然中鼩鼱的带毒率为50%,但由于其数量太少(2只),没有统计学意义。汉坦病毒病原体检出率较高,与以往调查结果相同[9-11]。
通过本次调查发现,以往大林姬鼠检出的汉坦病毒多为HTN型,而本次大林姬鼠检出的汉坦病毒均为SEO型,有待下一步研究。鼠种类分布及携带病原体情况见表2和图1。
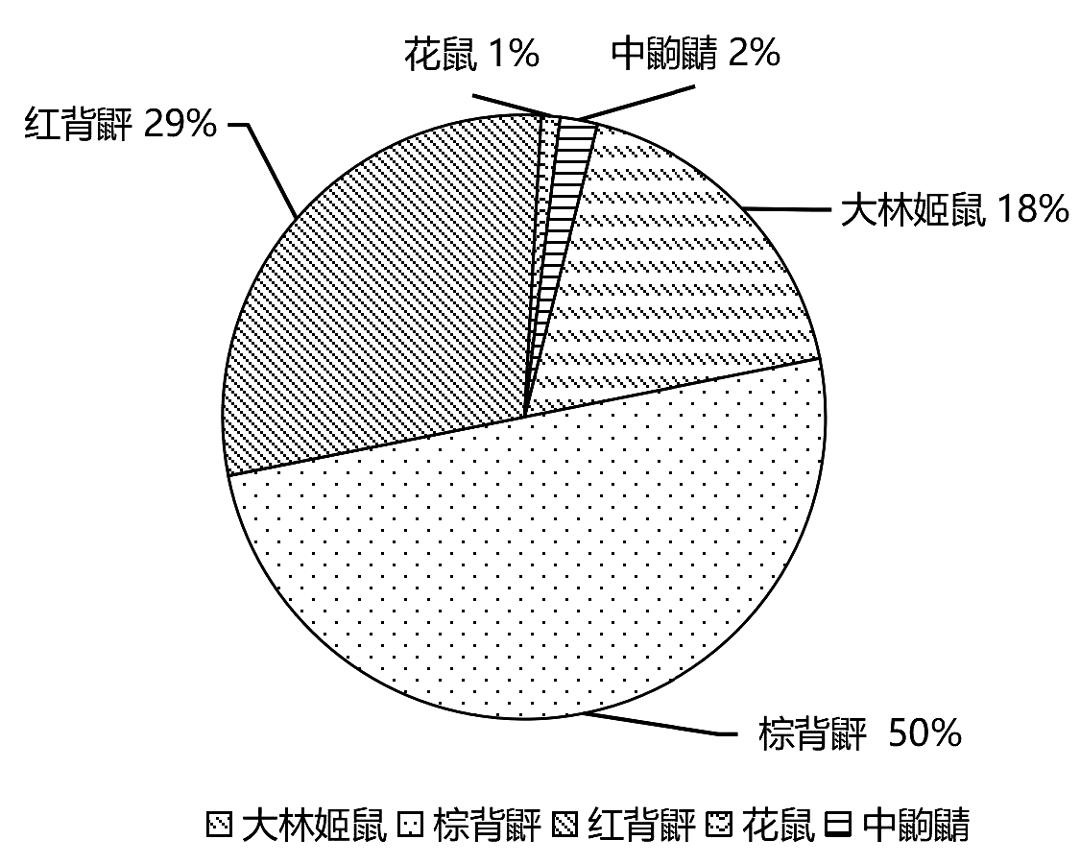
图1 双目峰口岸鼠种分布图
Fig.1 Distribution map of rat species at Shuangmufeng Port
表2 双目峰口岸鼠类汉坦病毒携带情况
Table 2 Hantavirus carriage in rodents at Shuangmufeng Port
种类 | (只) | (SEO) | (%) |
大林姬鼠 | 17 | 5 | 29.42 |
红背䶄 | 28 | 8 | 28.57 |
棕背䶄 | 49 | 8 | 16.33 |
花鼠 | 1 | 0 | 0 |
中鼩鼱 | 2 | 1 | 50.00 |
合计 | 97 | 22 | 22.68 |
2.2 汉坦病毒进化树的构建
从表3中汉坦病毒的信息可以看出汉坦病毒的基因编号、来源和发现地域各不同,说明各地病媒生物携带的汉坦病毒种类具有地域性差异。
使用MEGA 7.0软件,基于NJ法构建的M段汉坦病毒系统发育树显示,22个SEO型汉坦病毒测序样本都与朝鲜汉坦病毒DPRK08聚类在一起,也与NCBI-blast的比对结果一致(同源性在94%~99%),见表3和图2。
在本研究中,在双目峰口岸地区捕获的鼠共计5种97只,其中棕背䶄占50%、红背䶄占29%、大林姬鼠占18%、中鼩鼱2%、花鼠占1%,结果显示优势种为棕背䶄、红背䶄和大林姬鼠。以上结果均与以往鼠种调查结果相同[12-14]。
表3 参考汉坦病毒信息
Table 3 Reference to Hantavirus information
汉坦病毒 | 基因编号 | 病毒分型 | 来源 |
DPRK08 | JX853576.1 | SEO | North Korea |
EU163437.1 | SEO | China | |
XiaotangshanRn7 | GU592929.1 | SEO | China北京 |
Gongzhuling97 G1/G2 | KF745936.1 | SEO | China吉林 |
LYON/Rn/FRA/2013/LYO852 | KF387724.1 | SEO | France |
BjHD01 | DQ133505.1 | SEO | China 北京 |
Gongzhuling415 G1/G2 | KF745939.1 | SEO | China吉林 |
HBQ43 | KM233657.1 | SEO | China 河北 |
HuludaoRn101 | GU592923.1 | SEO | China 辽宁 |
Cherwell | KM948593.1 | SEO | England and Wales |
WuhanMm13 | JQ665886.1 | SEO | China 湖北 |
JiangxiXinjianRn-09-2011 | KP859514.1 | SEO | China 江西 |
A9 | AF035831.1 | HTN | China 湖北 |
HubeiHu02 | JQ665881.1 | HTN | China 湖北 |

图2 基于NJ法构建的部分M段汉坦病毒进化树
Fig.2 Partial M-segment Hantavirus evolutionary tree constructed based on NJ method
3 讨论
双目峰口岸是中朝两国边界线上唯一的陆路接壤通道,该口岸所在地我国吉林省安图县与朝鲜三池渊郡接壤,国境线长33.7 km,双目峰通道距长白山北坡山门23 km,距朝鲜三池渊郡35 km,是长白山地区重要的交通通道。2018年5月,国家口岸办批准安图双目峰公务通道作为临时口岸对外开放。双目峰口岸鼠类汉坦病毒流行状况尚未查到相关记载。
鼠是重要的病媒生物之一,可以携带百余种病原体和寄生虫。鼠携带的致病性汉坦病毒具有持续性和遗传性,并可通过其尿液、粪便和唾液产生的气溶胶、抓伤和咬伤等方式传播给人类[15]。每年在亚洲和欧洲发生数千例HFRS病例,其中大多数是由于感染HTNV、SEOV、PUUV或DOBV病毒。汉坦病毒感染目前在我国是较为严重的公共卫生问题之一,而且汉坦病毒感染的有效疫苗和治疗药剂难以开发和评价[16],因此汉坦病毒的监测和预防极其重要。
基于邻接法(NJ)构建的部分M段汉坦病毒进化树显示,22例阳性样本全与朝鲜汉坦病毒DPRK08聚类在一起[17],与吉林省Gongzhuling97 G1/G2汉坦病毒分为两支。表明双目峰地区汉坦病毒与朝鲜汉坦病毒同源性更高,与NCBI-blast比对结果一致(同源性94%~99%)。
我国与朝鲜有着较长的国境线,朝鲜地区病媒生物监测资料较少,据现有资料显示,朝鲜也是HFRS的流行区之一,因此充分了解中朝毗邻区域汉坦病毒在啮齿动物中的流行态势和扩散传播趋势,能有效地促进两国边境地区汉坦病毒的防控。目前双目峰口岸地区虽然尚未有人居住,但随着中朝两国关系的发展,未来双目峰口岸可能成为中朝两国重要的往来枢纽;另外,海关关员及边防警察、武警战士等也在边境线附近工作和生活,因此应积极改进和加强该地区病媒生物监测和预防控制措施,减少汉坦病毒等病媒传染病的发病率,更好地保障我国人民的生命安全。
参考文献
[1] Hong Jiang, Xuyang Zheng, Limei Wang, et al. Hantavirus infection: a global zoonotic challenge[J]. Virologica Sinica, 2017, 32(1): 32-43.
[2] H W Lee, P W Lee, K M Johnson. Isolation of the etiologic agent of Korean Hemorrhagic fever[J]. The Journal of infectious diseases, 1978, 137(3): 298-308.
[3]张海林, 张云智.中国汉坦病毒基因型及分布[J]. 中国媒介生物学及控制杂志, 2011, 22(5): 417-420.
[4]张吉, 刘勇先, 姚李四, 等. 2014-2015年吉林长白口岸鼠类监测结果分析[J]. 中国国境卫生检疫杂志, 2015, 38(11): 7-9.
[5] Kumar Sudhir, Stecher Glen, Tamura Kochiro. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets[J]. Molecular Biology and Evolution, 2016, 33(7): 1870-1874.
[6] Wei Zhang, Zhirong Sun. Random local neighbor joining: A new method for reconstructing phylogenetic trees[J]. Molecular Phylogenetics and Evolution, 2008, 47(1): 117-128.
[7]刘师文, 张艳妮, 肖芳, 等. 江西省高安市两株汉城病毒分离鉴定及全基因组序列分析[J]. 中国人兽共患病学报, 2022, 38(2): 102-107.
[8]范胜涛, 高晓龙, 李元果, 等. 吉林省汉坦病毒宿主动物携带病毒的遗传进化分析[J]. 中国生物制品学杂志, 2014, 27(4): 467-475.
[9]华满堂, 金兆清, 何成, 等. 新疆哈巴河铁热克提林牧区棕背䶄携带伯氏疏螺旋体的研究[J]. 中国人兽共患病学报, 2000, 16(3): 110.
[10]林革, 邹晓龙, 孙佳, 等. 双目峰口岸鼠类分子鉴定及进化分析[J]. 中国国境卫生检疫杂志, 2020, 43(1): 29-31.
[11] Li-Si Yao, Chen- Feng Qin, Yun Pu, et al. Complete genome sequence of Seoul virus isolated from Rattus norvegicus in the Democratic People’s Republic of Korea[J]. Journal of Virology, 2012, 86(24): 13853.
[12]金建丽, 刘铸, 田新民, 等. 东北地区棕背䶄种群遗传结构及亚种分类[J]. 兽类学报, 2013, 33(4): 383-388.
[13]李彤, 刘培琦, 姜岚. 棕背、红背䶄栖息地选择的研究[J]. 兽类学报, 1991, 11(1): 73-74.
[14]马杰, 李庆芬, 孙儒泳, 等. 东灵山辽东栎林啮齿动物群落组成及优势种大林姬鼠的繁殖特征[J]. 动物学报, 2003(2): 262-265.
[15]汪娜, 尹家祥. 肾综合征出血热流行过程及其影响因素研究进展[J]. 中国血吸虫病防治杂志, 2022, 34(2): 200-203.
[16] Papa A, Vaheri A, LeDuc J W, et al. Meeting report: Tenth International Conference on Hantaviruses[J]. Antiviral Research, 2016, 1(133): 234–241.
[17]张文义, 江佳富, 姚昆, 等. 中国大林姬鼠携带Amur类汉坦病毒及其分子生物学特征分析[J]. 中华流行病学杂志, 2007, 28(5): 482-486.
表1 汉坦病毒病原检测引物
Table 1 Pathogen detection primers
名称 | 引物 | 序列 (5'-3' ) | 位置 | 片段 (bp) |
逆转录 | P14 | TAGTAGTAGACTCC | ||
通用型 | HTY-F | AAAGTAGGTGITAYATCYTIACAATGTGG | 1910~1939 | M (+) |
HTY-R | GTACAICCTGTRCCIACCCC | 2373~2354 | M (-) | |
分型特异性引物 | ||||
汉滩型 | HTN-MF | GAATCGATACTGTGGGCTGCAAGTGC | 1958~1984 | M (+) |
HTN-MR | GGATTAGAACCCCAGCTCGTCTC | 2318~2340 | M (-) | |
汉城型 | SEO-MF | GTGGACTCTTCTTCTCATTATT | 1936~1957 | M (+) |
SEO-MR | TGGGCAATCTGGGGGGTTGCATG | 2331~2353 | M (-) | |
.
