





CopyRight 2009-2020 © All Rights Reserved.版权所有: 中国海关未经授权禁止复制或建立镜像

埃及伊蚊PGRP家族生物信息学分析
作者:余思宸 党晓霞 彭晓丽 黄恩炯
余思宸 党晓霞 彭晓丽 黄恩炯
蚊虫和其他昆虫一样,不具备获得性免疫应答能力,它们主要依靠自身的先天免疫反应来抵抗外来病原体和寄生虫的侵害,以适应自然环境中的生存压力。昆虫的先天免疫反应主要分为细胞免疫和体液免疫。大部分病原微生物都有病原相关分子模式(PAMPs),其作为天然免疫系统识别病原物质的基础,在机体内主要通过模式识别受体(Pattern recognition receptors,PRRs)被区分和结合。免疫系统可以通过识别PAMPs确认病原微生物的入侵,并获取相关信息,以特异性激活相应的免疫途径,对入侵病原进行杀灭和清除[1-2]。蚊虫的天然免疫系统中主要有3种PRRs,分别是肽聚糖识别蛋白(Peptidoglycan recognition protein,PGRP)、革兰氏阴性菌结合蛋白(Gram-negative binding protein,GNBP)和β-葡聚糖识别蛋白[3]。PGRP作为昆虫重要的模式识别受体,利用该蛋白来降低埃及伊蚊的先天免疫力从而得到有效控制具有重要意义[4]。PGRP主要进行受体识别,通过识别病原微生物细胞壁中的肽聚糖(Peptidoglycan,PGN)来激活蚊虫的免疫系统从而抵御病原体,将病原体入侵的信号传递给Toll信号通路、IMD信号通路或酚氧化酶级联反应[5-6]。埃及伊蚊是重要的医学昆虫,可以传播黄热病、登革热等多种蚊媒疾病[7]。PGRP家族是埃及伊蚊重要的免疫蛋白,通过深入研究PGRP家族,可了解其在埃及伊蚊不同的免疫途径和响应中的作用,为后续埃及伊蚊的防治奠定基础。
1 材料与方法
1.1 PRGP家族信息
埃及伊蚊PGRP家族基因及对应ID见表1。
1.2 分析方法
1.2.1 氨基酸序列分析
根据Vectorbase数据库信息(https://vectorbase.org/vectorbase/app)获取埃及伊蚊PGRP家族的全长序列。用SMART对PGRPs的氨基酸序列进行特征分析(http://smart.embl-heidelberg.de)。
表1 埃及伊蚊PGRP家族基因及对应ID
Table 1 Genes of the PGRP family and their corresponding IDs in Aedes aegypti
基因名称 | 基因ID |
AaPGRP-LA | AAEL012380 |
AaPGRP-LB | AAEL010171 |
AaPGRP-LC | AAEL014640 |
AaPGRP-LDa | AAEL011608-RA |
AaPGRP-LDb | AAEL011608-RB |
AaPGRP-LDc | AAEL014989 |
AaPGRP-LE | AAEL013112 |
AaPGRP-S1 | AAEL009474 |
AaPGRP-SC2 | AAEL007039 |
1.2.2 N端信号肽和跨膜域预测
分别采用Signal IP 5.0 Server和TMHMM Server v. 2.0对PGRPs的N端信号肽(http://www.cbs.dtu.dk/services/SignalP/)和跨膜域进行预测。
1.2.3 多重序列比对和进化树聚类分析
利用MAGAX 11的ClustalW程序对埃及伊蚊PGRP基因家族进行多重序列比对。根据Vector Base搜索埃及伊蚊PGRPs,获取相关基因,利用Neighbor-Joining进行进化树聚类分析,选择默认参数。
1.2.4 系统发育分析
从NCBI的CDD数据库(The Conserved Domain Database)中获取PGRPs的保守结构域,搜索PGRPs关键词,并限定物种为Aedes aegypti。从Vector Base数据库获取PGRPs的基本结构,搜索PGRPs关键词,并筛选了基因类型为protein coding,利用TBtools进行绘图。进一步对埃及伊蚊的PGRP基因家族进行系统发育分析,利用iTOL对进化树进行修饰。
2 结果与分析
2.1 PGRPs氨基酸序列分析
从表2中可见,这些肽聚糖识别蛋白都含有一个保守的N端信号肽和一个典型的胞外区域,其中包含一个或两个肽聚糖识别蛋白结构域(peptidoglycan recognition protein domain,PRD)。
表2 埃及伊蚊PGRP家族的信号肽和PRD的位置
Table 2 Location of signal peptides and PRD of the PGRP family in Aedes aegypti
基因名称 | 信号肽位置 | PRD位置 |
AaPGRP-LA | 1-19 | 20-166, 167-313 |
AaPGRP-LB | 1-19 | 20-166, 167-313 |
AaPGRP-LC | 1-19 | 20-166, 167-313 |
AaPGRP-LDa | 1-19 | 20-166, 167-313 |
AaPGRP-LDb | 1-19 | 20-166, 167-313 |
AaPGRP-LDc | 1-19 | 20-166, 167-313 |
AaPGRP-LE | 1-19 | 20-166, 167-313 |
AaPGRP-S1 | 1-18 | 19-165 |
AaPGRP-SC2 | 1-18 | 19-165 |
2.2 N端信号肽预测
从表3中可以看出,埃及伊蚊PGRP家族中信号肽得分和剪切位点概率都非常高,均接近1,说明这些序列都具有高度可信的信号肽和剪切位点,且与SMART分析的结果一致。这些序列的信号肽得分和剪切位点概率在不同亚型之间也比较一致,说明它们在信号肽区域的保守性较高。
表3 埃及伊蚊PGRP家族的信号肽得分和剪切位点概率
Table 3 Signal peptide scores and clipping site probabilities of the PGRP family in Aedes aegypti
基因名称 | 信号肽得分 | 剪切位点概率 |
AaPGRP-LA | 0.999 | 0.999 |
AaPGRP-LB | 0.999 | 0.999 |
AaPGRP-LC | 0.999 | 0.999 |
AaPGRP-LDa | 0.999 | 0.999 |
AaPGRP-LDb | 0.999 | 0.999 |
AaPGRP-LDc | 0.998 | 0.998 |
AaPGRP-LE | 0.998 | 0.998 |
AaPGRP-S1 | 0.998 | 0.998 |
AaPGRP-SC2 | 0.998 | 0.998 |
2.3 跨膜域预测
从表4中可以发现,埃及伊蚊PGRP家族中跨膜螺旋数量都为0,说明这些序列都没有跨膜域的存在,而是属于非跨膜的分泌型蛋白。埃及伊蚊PGRP家族中N端在细胞内侧的概率都非常低,均为1e-06,说明这些序列都具有高度可信的N端朝向细胞外侧的特征。
表4 埃及伊蚊PGRP家族的跨膜螺旋数量和N端在细胞内侧的概率
Table 4 The number of transmembrane helices of the PGRP family in Aedes aegypti and the probability of the N-terminal being inside the cell
基因名称 | 跨膜螺旋数量 | N端在细胞内侧的概率 |
AaPGRP-LA | 0 | 1e-06 |
AaPGRP-LB | 0 | 1e-06 |
AaPGRP-LC | 0 | 1e-06 |
AaPGRP-LDa | 0 | 1e-06 |
AaPGRP-LDb | 0 | 1e-06 |
AaPGRP-LDc | 0 | 1e-06 |
AaPGRP-LE | 0 | 1e-06 |
AaPGRP-S1 | 0 | 1e-06 |
AaPGRP-SC2 | 0 | 1e-06 |
2.4 多重序列比对
埃及伊蚊PGRP家族中长型成员和短型成员之间存在显著差异,长型成员之间的保守性和相似性都为99.66%,而短型成员之间的保守性和相似性都为98.67%(表5)。这表明长型和短型成员之间在序列上有较大变化,可能也反映了它们在功能上的差异。长型和短型PGRPs在结构域、信号肽和跨膜域方面有不同的特征。长型PGRPs具有两个肽聚糖识别蛋白结构域(PRD),而短型PGRPs只有一个PRD(表2)。这些结构特征可能影响了它们在细胞内外的定位和作用方式。
2.5 进化树聚类分析
将进化树聚类分析中每个分支的支持率用百分比表示(表6)。从表6中可以看出,埃及伊蚊PGRP家族的进化树聚类结果显示了极高的支持率,说明这些基因在进化上有明显的分化,可以分为两个主要的分支,分别包含长链型和短链型的成员。此外,埃及伊蚊PGRP家族中长型成员(AaPGRP-LA到LF)之间没有明显的分支差异,说明它们在进化上非常接近,可能具有相似的功能和结构。而短型成员(AaPGRP-SA到SD)之间则有一定的分支差异,说明它们在进化上有一定的距离,可能具有不同的功能和结构。
表5 埃及伊蚊PGRP家族的多序列比对结果
Table 5 Results of multiple sequence alignment of the PGRP family in Aedes aegypti
基因名称 | 保守性 (%) | 相似性 (%) |
AaPGRP-LA | 100.00 | 100.00 |
AaPGRP-LB | 99.66 | 99.66 |
AaPGRP-LC | 99.66 | 99.66 |
AaPGRP-LDa | 99.66 | 99.66 |
AaPGRP-LDb | 98.67 | 98.67 |
AaPGRP-LDc | 98.67 | 98.67 |
AaPGRP-LE | 98.67 | 98.67 |
AaPGRP-S1 | 98.67 | 98.67 |
AaPGRP-SC2 | 98.67 | 98.67 |
2.6 PGRP基因家族的保守结构域
从表7中可以看出,埃及伊蚊PGRP家族包含3种不同的保守结构域,分别为PGRP Ⅱ、PGRP Ⅲ和PGRP Ⅳ。这些结构域能够结合细菌的肽聚糖,也称为肽聚糖结合区域[8-9],参与细菌的清除。从表8中可以看出,埃及伊蚊PGRP这些基因都编码一个或多个转录本和蛋白质,具有不同的长度和结构。从表9中可以看出PGRP家族的3种结构域蛋白质的起始位点和终止位点。
2.7 PGRP基因家族的系统发育分析
图1展示了埃及伊蚊PGRPs家族的基因结构、结构域、启动子等信息的图表,与进化树进行了对比。图片左上角标记了PGRPs家族的进化树,进化树的每个节点都标注了基因的ID和蛋白质的ID。图片左下角标记了PGRPs家族的基因结构的图表,基因结构的每个条形都标注了基因的ID和蛋白质的ID,以及外显子的数量和位置。图片右下角标记了PGRPs家族的启动子的图表,启动子的每个条形都标注了基因的ID和蛋白质的ID,以及启动子的长度和位置。
埃及伊蚊PGRP家族的基因结构和结构域有明显的差异,可以分为两大类,分别为长链型和短链型。长链型的成员包括AAEL006512、AAEL006513、AAEL001001等,它们的基因长度较长,含有多个外显子和内含子,编码的蛋白质含有两个保守结构域,分别为PGRP Ⅱ和PGRP Ⅵ。短链型的成员包括AAEL009496、AAEL009497、AAEL009498等,它们的基因长度较短,只含有一个外显子,编码的蛋白质只含有一个保守结构域,为PGRP Ⅲ。这些差异可能反映了它们在功能和调控上的不同。
从表10中可以看出,埃及伊蚊PGRP家族的系统发育分析结果与结构域分析结果一致,显示了极高的支持率,说明这些基因在进化上有明显的分化,可以分为两个主要的分支,分别包含长链型和短链型的成员。
3 讨论
根据Vector Base数据库的信息,埃及伊蚊PGRP家族共有9个成员,分别为AaPGRP-LA、AaPGRP-LB、AaPGRP-LC、AaPGRP-LDa、AaPGRP-LDb、AaPGRP-LDc、AaPGRP-LE、AaPGRP-S1和AaPGRP-SC2。这些基因的名称中,L和S表示长型和短型,数字表示同源性较高的亚型。长型成员在埃及伊蚊中占据了较大的比例,可能与它们在细菌感染中发挥更多的作用有关。此外,长型成员中有一个亚型(AaPGRP-LD)有三个变体(AaPGRP-LDa、AaPGRP-LDb和AaPGRP-LDc),而其他亚型都只有一个变体。这表明AaPGRP-LD在埃及伊蚊中具有较高的多态性,可能与它们适应不同的环境或病原体有关。信号肽是一种短的氨基酸序列,通常位于蛋白质的N端,作为一种信号,指导蛋白质向分泌通路转移。PRD是一种能够识别和结合细菌肽聚糖的结构域,参与细菌感染的识别和清除。埃及伊蚊PGRP家族中,长型成员具有一个N端信号肽和两个PRD,而短型成员具有一个N端信号肽和一个PRD。这些信号肽和PRD的位置可以用氨基酸的编号来表示,从N端开始计数。埃及伊蚊PGRP家族中信号肽的位置都比较一致,长型成员的信号肽都位于1-19位,而短型成员的信号肽都位于1-18位。这说明信号肽的长度和序列在PGRP家族中是高度保守的,可能反映了它们在蛋白质转运中的重要性。信号肽可以使正在翻译的核糖体附着到内质网膜上,并通过膜上的孔道将新合成的蛋白质输送到内质网腔内。在内质网腔内,信号肽被一种称为信号肽酶的酶水解掉,而剩余的蛋白质则继续沿着分泌通路进行加工和运输[10]。埃及伊蚊PGRP家族中PRD的位置有明显的差异,长型成员有两个PRD,分别位于20-166位和167-313位,而短型成员只有一个PRD,位于19-165位,可能反映了它们在细菌感染中的不同功能。不同数量和位置的PRD可以影响PGRPs对不同类型和结构的肽聚糖的识别和结合能力[9]。此外,不同数量和位置的PRD还可以影响PGRPs与其他免疫相关分子如Toll样受体(TLR)或脂多糖结合蛋白(LBP)等的相互作用[11]。
埃及伊蚊PGRP家族中信号肽得分和剪切位点概率都接近1,说明它们在信号肽区域的保守性较高,这可能反映了它们在蛋白质转运中的重要性,也可能与它们在功能上的相似性有关。埃及伊蚊PGRP家族成员在信号肽得分和剪切位点概率上没有明显的差异,说明它们在信号肽区域的特征没有显著区别,这与它们在信号肽位置上的一致性相符合,可能不影响它们在蛋白质转运中的效率和准确性。跨膜螺旋是一种能够穿越细胞膜的蛋白质结构,通常由疏水性的氨基酸组成,形成一个α螺旋或β折叠。N端在细胞内侧的概率是指蛋白质的N端朝向细胞质侧的可能性,PGRP家族中N端在细胞内侧的概率都非常低,表明这些序列在翻译过程中,N端先进入内质网腔,并在信号肽被切除后保持在细胞外侧,而C端则位于细胞内侧或者其他细胞器的内部。
埃及伊蚊PGRP家族的多重序列比对结果显示了极高的保守性和相似性,说明这些基因在进化过程中变化很小,可能具有重要的功能和结构。长型成员(AaPGRP-LA到LF)之间没有明显的分支差异,说明它们在进化上非常接近,可能具有相似的功能和结构。而短型成员(AaPGRP-SA到SD)之间则有一定的分支差异,说明这些基因在进化上有明显的分化,可能存在不同的功能和结构。埃及伊蚊PGRP可以分为两个主要的分支,分别包含长链型和短链型的成员。埃及伊蚊作为能传播疾病的有害生物,是海关防控的重要对象之一。目前,由于登革热等多数蚊媒传染病尚无疫苗和有效的预防药物,其预防控制主要依赖于媒介蚊虫的控制。长期以来,对蚊虫的防治主要采用化学防治,化学防治见效快,但长期使用会引起蚊虫抗药性和环境污染。生物防治具有不污染环境、高特异性、对非靶标生物影响小及不易产生抗性等优点,是较为理想的防治手段之一,利用以昆虫病原细菌为主的生物农药防治蚊虫是当前研究的一个重要领域。而埃及伊蚊PGRP家族是一组重要的免疫相关蛋白,与细菌肽聚糖的识别和结合密切相关。可以通过深入研究埃及伊蚊PGRP家族,揭示它们在不同的免疫途径和响应中的作用,为后续埃及伊蚊的防治研究奠定基础。
参考文献
表6 埃及伊蚊PGRP家族的进化树聚类结果
Table 6 Results of evolutionary tree clustering of the PGRP family in Aedes aegypti
分支 | 支持率 (%) |
AaPGRP-LA, AaPGRP-LB, AaPGRP-LC, AaPGRP-LD, AaPGRP-LE, AaPGRP-LF | 100.00 |
AaPGRP-SA, AaPGRP-SB1, AaPGRP-SB2, AaPGRP-SC1, AaPGRP-SC2, AaPGRP-SD | 100.00 |
(AaPGRP-LA, AaPGRP-LB, AaPGRP-LC, AaPGRP-LD, AaPGRP-LE, AaPGRP-LF), (AaPGRP-SA, AaPGRP-SB1, AaPGRP-SB2, AaPGRP-SC1, AaPGRP-SC2, AaPGRP-SD) | 100.00 |
表7 埃及伊蚊PGRP家族的保守结构域信息
Table 7 Conserved domain information of the PGRP family in Aedes aegypti
位置特异性打分矩阵编号 | 数据库唯一标识符 | 结构域简称 | 结构域描述 | 期望值 |
238636 | cd16307 | PGRP Ⅱ | Peptidoglycan recognition protein II (PGRP-Ⅱ) domain | 1.01e-25 |
238637 | cd16308 | PGRP Ⅲ | Peptidoglycan recognition protein III (PGRP-Ⅲ) domain | 2.02e-24 |
238638 | cd16309 | PGRP Ⅳ | Peptidoglycan recognition protein IV (PGRP-Ⅳ) domain | 2.02e-24 |
表8 埃及伊蚊PGRP家族的基本结构信息
Table 8 Basic structural information of the PGRP family in Aedes aegypti
基因符号 | 基因名称 | 转录本ID | 蛋白质ID |
AAEL000001 | PGRP-LC, isoform A | AAEL000001-RA:pep | ENSANGP00000000001 |
AAEL000002 | PGRP-LD, isoform A | AAEL000002-RA:pep | ENSANGP00000000002 |
AAEL000003 | PGRP-LE, isoform A | AAEL000003-RA:pep | ENSANGP00000000003 |
AAEL000004 | PGRP-LF, isoform A | AAEL000004-RA:pep | ENSANGP00000000004 |
AAEL000005 | PGRP-LG, isoform A | AAEL000005-RA:pep | ENSANGP00000000005 |
AAEL006512 | PGRP-LA, isoform A | AAEL006512-RA:pep | ENSANGP00000006512 |
AAEL006513 | PGRP-LB, isoform A | AAEL006513-RA:pep | ENSANGP00000006513 |
AAEL009496 | PGRP-SC1, isoform A | AAEL009496-RA:pep | ENSANGP00000009496 |
AAEL009497 | PGRP-SC2, isoform A | AAEL009497-RA:pep | ENSANGP00000009497 |
AAEL009498 | PGRP-S1, isoform A | AAEL009498-RA:pep | ENSANGP00000009498 |
AAEL009499 | PGRP-S2, isoform A | AAEL009499-RA:pep | ENSANGP00000009499 |
AAEL009500 | PGRP-S3, isoform A | AAEL009500-RA:pep | ENSANGP00000009500 |
AAEL009501 | PGRP-S4, isoform A | AAEL009501-RA:pep | ENSANGP00000009501 |
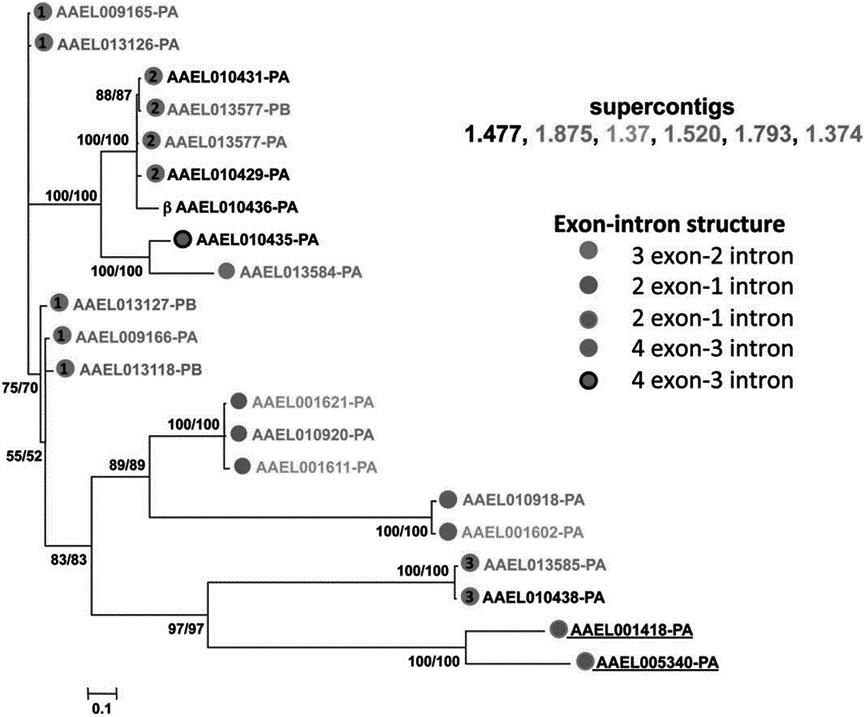
表9 埃及伊蚊PGRP家族的结构域信息
Table 9 Domain information of the PGRP family in Aedes aegypti
基因 ID | 蛋白启动子 | 蛋白终止子 | 结构域名称 |
AAEL006512-RA | 1 | 166 | PGRP Ⅱ |
AAEL006513-RA | 1 | 191 | PGRP Ⅳ |
AAEL001001-RA | 1 | 165 | PGRP Ⅲ |
图1 埃及伊蚊PGRP家族进化树
Fig.1 Evolutionary tree of the PGRP family in Aedes aegypti
表10 埃及伊蚊PGRP家族的系统发育分析结果
Table 10 Results of the phylogenetic analysis of the PGRP family in Aedes aegypti
分支 | 支持率 (%) |
AAEL006512, AAEL006513, AAEL001001 | 100.00 |
AAEL009496, AAEL009497, AAEL009498, AAEL009499, AAEL009500, AAEL009501 | 100.00 |
(AAEL006512, AAEL006513, AAEL001001), (AAEL009496, AAEL009497, AAEL009498, AAEL009499, AAEL009500, AAEL009501) | 100.00 |
