





CopyRight 2009-2020 © All Rights Reserved.版权所有: 中国海关未经授权禁止复制或建立镜像

光肩星天牛与星天牛地理分布及生态限制因子研究
作者:彭梦香 莫瑾 寇鑫 梁新苗 严珺
彭梦香 莫瑾 寇鑫 梁新苗 严珺
彭梦香 1 莫 瑾 1 寇 鑫 2 梁新苗 2 * 严 珺 1
摘 要 光肩星天牛和星天牛是为害杨属、柳属、槭属、榆属等植物的重要林木蛀干害虫,其分布受多种生态环境因素的影响。近年来,光肩星天牛有向高纬度、高海拔扩散的潜在风险。因此,明确影响两种天牛分布的生态因子,对于预测其潜在适生区、制定早期预警机制和精准防控策略具有重要意义。本文针对温度、海拔、寄主、天敌等关键生态限制因子及其在国内的分布情况进行了分析,并对两种天牛的防治策略进行了探讨,为保障中国木制品出口以及加强林业有害生物风险管理提供参考依据。
关键词 光肩星天牛;星天牛;分布;生态因子
Study on Geographical Distribution and Ecological Limiting Factors of Anoplophora glabripennis and Anoplophora chinensis
PENG Meng-Xiang 1 MO Jin 1 KOU Xin 2 LIANG Xin-Miao 2* YAN Jun 1
Abstract Anoplophora glabripennis and Anoplophora chinensis are important wood-boring pests that infest trees such as Populus, Salix, Acer and Ulmus. They are both listed as worldwide quarantine pests, and their distribution is affected by various ecological and environmental factors. In recent years, Anoplophora glabripennis has a potential risk of spreading to higher latitudes and altitudes. Therefore, identifying the ecological factors affecting the distribution of these two pests is of great significance for predicting their potential suitable habitats, establishing early warning mechanisms, and formulating precise prevention and control strategies. This paper analyzes the key ecological limiting factors such as temperature, altitude, host plants and natural enemies, as well as their distribution within China. It also discusses control strategies for both pests, providing valuable references for safeguarding China’s wood product exports and managing forest pest risks.
Keywords Anoplophora glabripennis; Anoplophora chinensis; distribution; ecological factors
本论文由水稻种子品质检测及健康安全技术1+X研究室资助
第一作者:彭梦香(1994—),女,土家族,湖南龙山人,硕士,中级工程师,主要从事植物检验工作,E-mail: 272373871@qq.com
通信作者:梁新苗(1976—),女,汉族,新疆乌鲁木齐人,博士,研究员,主要从事植物检验及生物安全研究工作, E-mail: hellosun21@163.com
1. 长沙海关技术中心 长沙 410004
2. 中国海关科学技术研究中心 北京 100026
1. Technology Center of Changsha Customs, Changsha 410004
2. Science and Technology Research Center of China Customs, Beijing 100026
光肩星天牛和星天牛均属于鞘翅目、天牛科、星天牛属,是在中国分布较广、为害杨属(Populus)、槭属(Acer)、柳属(Salix)、榆属(Ulmus)等较多树种的一类杂食性林木蛀干昆虫,既蛀食树干,也为害嫩枝,幼期通过破坏树木的疏导组织,切断水分和养分的运输,导致树木衰弱,甚至死亡[1-4]。天牛幼虫生活隐蔽、蛀孔复杂,成虫体壁、鞘翅厚而坚硬,耐药力强,世代较长并参差不齐,繁殖速度快,防治难度大[5-6],给当地生态环境和林业经济带来严重威胁。在国际贸易中,这两种天牛主要通过种植苗木、木制包装等方式传播,20世纪90年代末因频繁在世界各海关被截获并传入欧洲而备受关注[7]。目前,光肩星天牛已在美国部分地区定殖,两种天牛均在欧洲多个国家和地区建立了种群[8],被欧洲和地中海植物保护组织(European and Mediterranean Plant Protection Organization,EPPO)列为重要林木检疫对象。
对于陆地生存的光肩星天牛和星天牛而言,其分布受多种生态环境因素的影响,诸如气候、地理环境、寄主树种以及自身适应性等[9]。Yasser Alramadan等[10]通过模型分析发现年平均温度、日较差、温度季节性变化及最湿润月份降雨量是影响星天牛栖息地适宜性的重要变量。KAPPEL A P等[11]将美国作为研究区域加以研究发现,温度较高、寄主物种丰富的地区光肩星天牛发生风险较大。国内研究发现植被覆盖率、郁闭度和林分位置对光肩星天牛的发生有显著影响,植被覆盖率和郁闭度越高,害虫数量越少[12]。针对天牛种群寄生与扩散的有关研究发现,光肩星天牛倾向于选择树干直径较大的树木寄生,成虫的扩散密度随距离和时间的增加而减少,扩散格局与风向有关[13-15]。近年来,中国光肩星天牛正在向高纬度高海拔地区扩散[16],新的发生区已北达哈尔滨、东达牡丹江、西达伊犁,高海拔至西藏等地区[17]。因此,明确影响两种天牛分布的生态因子,对于预测其潜在适生区、制定早期预警机制和精准防控策略具有重要意义。本文通过分析温度、湿度、海拔、寄主植物及天敌等生态因素对两种天牛地理分布的影响,探讨了两种天牛的防控措施,以期为应对技术性贸易措施、保障中国木制品出口贸易以及加强林业有害生物风险管理提供理论支撑。
1 生态地理分布
1.1 形态特征
光肩星天牛(英文名:Asian longhorned beetle,ALB)和星天牛(英文名:Citrus longhorned beetle,CLB)均属完全变态昆虫,一个世代需要经历卵、幼虫、蛹和成虫4个虫态,体表均为漆黑色,有光泽。光肩星天牛成虫体长约19~39 mm,鞘翅上分布有10~20个白色或黄色不规则斑块,鞘翅基部光滑。触角11节,雄性触角与体长的比例范围为1.6~2.5,雌性为1.2~1.8。卵长椭圆形,两端略凹陷,初产时乳白色,后逐渐变为黄褐色。幼虫头部呈棕色,胸节和腹节通常为乳白色,幼龄幼虫体长7~20 mm,老熟幼虫体长30~60 mm。蛹约30 mm,乳白至黄白色,裸蛹,触角在腹面可见且呈螺旋状。
星天牛成虫体长19~41 mm,鞘翅上有15~20个白斑,排成不规则的5横行,鞘翅基部密布黑色小颗粒。雄虫触角超过体长1倍,雌虫的触角稍长于体。卵约5 mm,呈长椭圆形,初产时为乳白色,快孵化时变成黄褐色。老熟幼虫体长45~67 mm,圆筒形,稍扁,淡黄白色,胸部肥大,前胸背板前方左右各有一黄褐色飞鸟形斑纹,后方有一块“凸”字形大斑纹,略隆起。蛹为裸蛹,长约27~38 mm,羽化前渐变为淡黄色至黑褐色,触角细长卷曲,体似成虫。星天牛和光肩星天牛的外部形态特征极为相似,主要通过鞘翅基部是否具有颗粒状凸起进行区分。
1.2 地理分布
1958—1991年,苏筱雨等[19]调查了中国东北、华北、华东、中南及西北、西南地区光肩星天牛分布情况,确定北起辽宁北部,向西到内蒙古高原东部,向南经承德、张家口再向西经呼和浩特到宁夏,然后沿青藏高原东侧的兰州、成都、乐山,再由此向南到云南楚雄,此线以南以东为中国光肩星天牛东部分布区。2000—2003年,王志刚[20]对东北三省57个市、县进行光肩星天牛调查,光肩星天牛分布区向东部和北部扩散了2个经纬度,向西扩散15个经度。到目前为止,光肩星天牛在中国29个省市有过分布记录[21],但主要以长江以北地区为主。
星天牛主要生活在温暖的南方,在浙江、湖北、湖南、广东、海南等22个省市有过分布记录[22]。目前星天牛在中国总体发生危害较轻,未见大面积暴发的灾难性报道。特别是河南以北地区极少分布或没有分布,鲜见危害报道。
2 影响分布的生态限制因子
2.1 温度和湿度
温湿度条件是决定天牛生存的关键因子,光肩星天牛和星天牛都需要足够的积温指标才能完成各龄期的生长发育(表1)。张树怀等[23]在实验条件下发现光肩星天牛卵、一龄幼虫、二龄幼虫、蛹的有效积温分别为250.3日度、157.8日度、232.0日度、182.6日度。杨志敏等[24]报道了光肩星天牛幼虫和蛹期以及全时代发育的有效积温分别为1231.89日度和1264.2日度。Keena M.A.[25]发现光肩星天牛的卵在10℃及以下很难孵化,而孵化的温度上限是35℃。预测光肩星天牛卵发育的下限温度为9.7℃,1 ~ 5龄幼虫和蛹期发育的下限温度在10℃左右,高龄幼虫在12℃左右。最高发育临界温度是35~40℃,在10~30℃范围内,发育历程跟温度呈线性关系[26]。陈辉等[27]提出最热月平均气温<15℃的地区达不到光肩星天牛化蛹的起点温度而使之不能生存。黄斑星天牛已被证实与光肩星天牛为同一种的不同型[28]。据周嘉熹等[29]研究,黄斑星天牛危害区的主要生态特点是冬季不很冷,夏季较凉爽,又具一定季节性高温期,同时具备黄斑星天牛取食树种,特别是感虫树种多,天敌少。夏季温度过高,超过32℃或夏季有效积温达不到化蛹要求的,黄斑星天牛很难猖獗发生,有分布而不成灾地区是气候因子的限制结果。仲丽等[30]发现高湿度条件下,光肩星天牛卵的孵化率比较高。王志刚[20]归纳出光肩星天牛分布区的年均温为3.9℃以上;春季均湿为37%以上,年均湿为39.4%以上,年降水量在130.4 mm 以上。危害严重地区的年均气温在3. 9℃以上,年均相对湿度44.9% 以上。
相比光肩星天牛,星天牛的高温适应能力更强,耐寒性更差。Melody等[31]研究发现成虫产卵的最高阈值温度、最低阈值温度和最适温度分别为35℃、10℃、29℃。预测星天牛卵的最低发育阈值温度为13℃,最高发育阈值温度为38℃,卵孵化的最适温度为26℃。当温度大于35℃时,雌虫不再产卵,成虫数量下降。蛹和1~6龄幼虫的发育最低阈值温度为10℃,高龄幼虫为12℃。Adachi[32]报道幼虫越冬后,需要累积1200°C的总温度才能从幼虫发育为成虫。黄金水等[33]室内实验发现蛹的发育起点温度为(13.91±0.69)℃,有效积温为(144.04±16.96)日度。星天牛分布的年均温在10℃以上、36℃以下[34]。
2.2 海拔
海拔通过调控氧分压、温度以及寄主植物资源等间接限制天牛分布范围。李文杰等[35]报道光肩星天牛在海拔1000 m以下都有分布,但是主要集中在海拔200 m以下。阎浚杰[36]通过对河北省6个地区的调查,归纳出光肩星天牛的垂直分布范围:海拔1000 m以上分布很少,是安全区;200~1000 m为偶发区;200 m以下分布最多。在中国境内,宁夏、甘肃和内蒙古等地区为光肩星天牛发生区,它们的海拔高度分别为1000~1200 m、1000 m以上和1000 m[37]。杨忠岐等[38]发现光肩星天牛在水平和垂直分布呈现一直向外和向上的趋势,有向高纬度或高海拔地区侵染的风险。王志刚等[39]报道西藏南部发生地点在海拔3000 m以上。这种向高海拔高纬度迁移的情况可能跟害虫本身的耐寒性以及全球气候变暖有关。光肩星天牛越冬状态的超冷却点是-2.4~-47.4℃[40]。韦美泽[17]发现光肩星天牛的耐低氧能力特别强,能够在氧气浓度很低的环境下生存,推测2%的氧浓度可能是该虫维持生命体征的耐受低限。全球气候变暖可能导致物种向北迁移[41]。在昆虫中,黑腹果蝇(Drosophila melanogaster)、绿蝽(Nezara viridula)和桔小实蝇(Bactrocera ordorsalis)已经被证实可以随着气候变暖向北迁移。Xuezhen Ge等[42]基于全球气候变暖预测未来适宜星天牛的分布范围为18.2~49.5°N和81.3~135.0°E,适生区未来向东北和西北方向扩张。BYEON等和Zhou等[43-44]研究表明,在30°N以北的地区,光肩星天牛的适生区将扩大,在30°N以南的大多数地区缩小。
2.3 寄主植物
光肩星天牛的寄主范围广,在全世界范围内已报道的寄主植物已有209种,包括10个目、21个科、41个属。光肩星天牛主要侵染阔叶树种,包括杨属(Populus)、槭属(Acer)、柳属(Salix)、榆属(Ulmus)、七叶树属(Aesculus)、桦属(Betula)等,大约75.12%的寄主植物种类属于这六个属,其中杨属占41.15%,槭属占12.92%,柳属占9.56%,榆属占5.74%,七叶树属占2.87%,桦属占2.87%(如图2)。约有86个杨属物种或品种容易受到损害,其中95.35%的杨属物种在中国有记录。光肩星天牛在不同地区表现出不同的寄主偏好,在中国,杨属和柳属是最常被侵染的树种,而槭属则是北美和欧洲最常被侵染的树种。据报道,中国、北美洲和欧洲地区分别有162种、36种和26种树木可作为光肩星天牛的寄主。研究人员以是否能够支持天牛完成整个生命周期为依据将寄主植物分为高感树种(High-sensitivity,HS)、中感树种(Moderate-sensitivity,MS)、部分敏感树种(Partial-sensitivity,PS)、低感树种(Low-sensitivity,LS)4类。研究结果显示,分别有101种、70种、11种和27种树木属于这四类。宿主植物种类在不同地区存在显著差异。在中国,分别有78种、63种、5种和17种树木被归为HS、MS、PS和LS类别;在北美洲地区,分别有16种、9种、7种和4种树木被归为HS、MS、PS和LS类别;在欧洲地区,分别有17种、4种、1种和4种树木被归为HS、MS、PS和LS类别[45]。
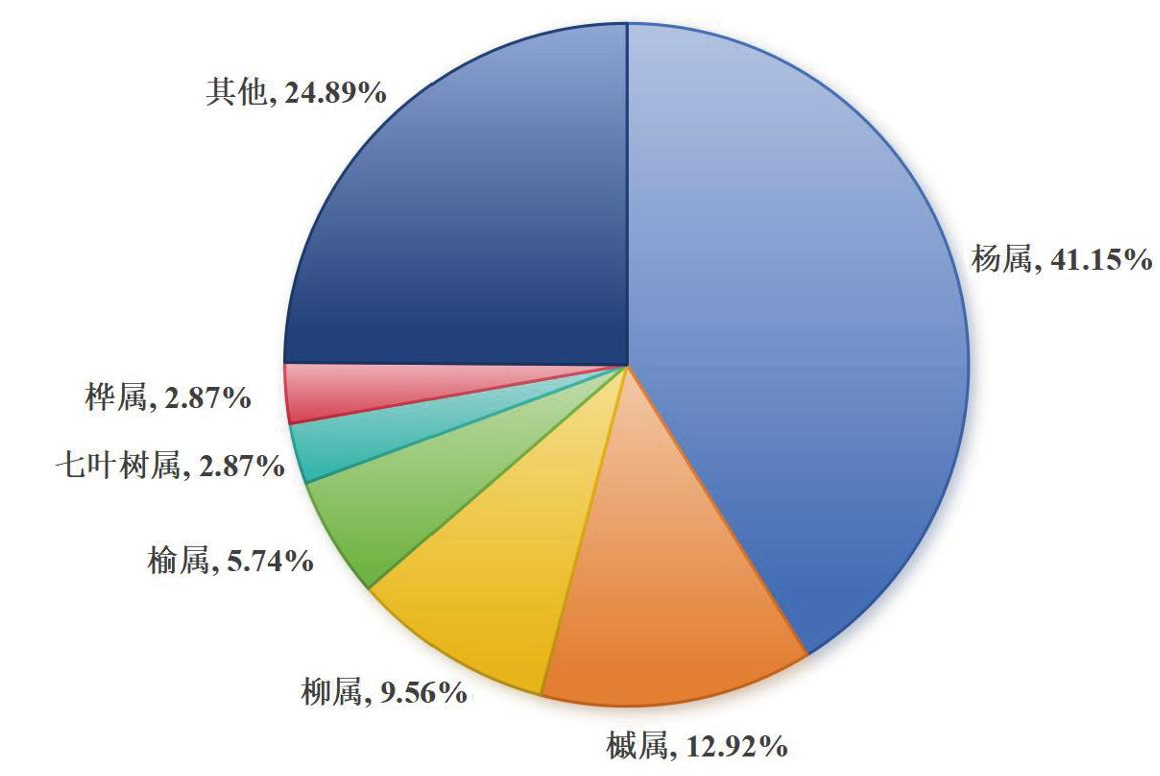
图2 光肩星天牛不同属寄主比例
Fig.2 Host proportion of different genera for Anoplophora glabripennis
星天牛危害的植物种类很多,我国最早报道的星天牛寄主植物是柑橘和木麻黄,后来在苹果、梨、无花果、樱桃、枇杷、花红、柳、白杨、桑、苦糠、柳豆、洋槐、榆、悬铃木、核桃、桑、红椿、花揪、乌桕、梧桐、相思树、楝、枣以及其他林果植物均有发现,共计29科29属[46]。在欧洲,不论是本土种还是外来种,星天牛喜食械类树种[47] 。截至目前,已报道的星天牛寄主植物在全世界范围内已超过100种[7],共38科[34]。但其中许多种类只有成虫补充营养阶段的取食记录,是否为寄主植物尚待证实。Henrik Sjöman等[48]根据星天牛是否在其上完成整个生活史或者是否有出虫孔,将植物分为优秀寄主、良好寄主、一般寄主以及抗性植物,归类优秀寄主6种,分别是槭属、柑橘属、苹果属、梨属、桦木属、榛属和紫薇属,良好寄主23种。
寄主树种对于物种分布特别是生活隐蔽、受环境条件波动性小的蛀干害虫的分布发挥着关键作用[48-50]。气候条件不适宜时,若有寄主植物存在,害虫也有可能定殖。Lee等[51]发现了光肩星天牛在韩国本土分布地二次扩散蔓延至韩国其他未分布区域的生态现象。国内学者对其他天牛类昆虫的实地调查和扩散研究也佐证了这一点[52]。在特定生态地理分布区内,寄主林木决定主要成灾的天牛种类,寄主林木丰富度则决定天牛种群数量,这是虫灾的基本特征[53]。
2.4 天敌
天敌在控制害虫种群数量方面发挥着重要作用,它们通过捕食、寄生或致病等方式抑制天牛的繁殖和迁移,减少向新区域扩散的风险,避免其在特定生态地理分布区域猖獗暴发。光肩星天牛的天敌有昆虫、鸟类和微生物3类[54]。鸟类主要是啄木鸟,如大斑啄木鸟(Dendrocopos major Malherbe)[55];微生物有白僵菌、苏云金杆菌等;昆虫有花绒穴甲(Dastarcus helophoroides Fairmaire)、管氏肿腿蜂(Sclerodermus guani Xiao et Wu )、白蜡吉丁肿腿蜂(Sclerodermus pupariae Yang et Yao)、柄腹茧蜂(Spathius anoplophorae)、赤腹茧蜂(Iphiaulax imposter Scopoli)、草地铺道蚁(Tetramorium caespitum L.)、膜毛绥螨(Lasioseius ometes Berlese)、毛绥螨(L.sp.)、蠹蛾肛厉螨(Proctolaelaps cossi Muler)、枝厉螨(Dendrolaelap ssp.)、奥金小蜂(Oxysychus sp.)等10余种[56-57],其中花绒寄甲是一种优势天敌,自然寄生率为 9.4%[54]。Duan J J等[58]在美国中大西洋地区的红枫上发现了寄生于光肩星天牛幼虫期的天敌,调查结果显示 5 组茧蜂科的寄生蜂能够成功寄生光肩星天牛小幼虫期,其中密足陡盾茧蜂(Ontsira mellipes Ashmead)被认为是一种可应用于光肩星天牛生物防治的潜在天敌。李飞[59]利用砍伐自然受害树解剖的方法调查了北京、天津、珲春三地的光肩星天牛自然天敌种类及作用贡献率,发现捕食天敌所占的比例最大,北京地区捕食率高达65.5%。目前肿腿蜂和花绒寄甲是光肩星天牛生物防治所用的主要天敌,肿腿蜂寄生光肩星天牛低龄幼虫期,花绒寄甲主要寄生光肩星天牛老熟幼虫期和蛹期。对北京市房山区和大兴区旱柳上进行花绒寄甲释放试验,房山区光肩星天牛排粪孔数量最终减少85.42%,幼虫的数量下降70%~82.5%;大兴区光肩星天牛排粪孔数量最终减少71.03%,幼虫数量下降57.14%~100%[60]。在甘肃省张掖市释放花绒寄甲成虫和卵卡后,花绒寄甲平均寄生率可达30.05%,防治效果最高为41.28%[61]。
国内外已发现许多可以寄生在星天牛各个生长发育时期起到控制作用的天敌种类,包括分布在中国、韩国以及日本等地的花绒寄甲以及亚洲和欧洲等地的一些拟寄生蜂种类[62]。国内记录的有:蚂蚁、蠼螋、川硬皮肿腿蜂(Scleroderma sichuanensis Xiao)、管氏肿腿蜂(S.guani Xiao et Wu)、天牛卵长尾啮小蜂(Aprostocetus. fukutai Miwa & Sonan)、两色刺足茧蜂(Zombrus bicolor Enderlein)、斑头陡盾茧蜂(Ontsira palliates Cameron)等。在欧洲本土发现有 9 种本地拟寄生蜂:欧洲硬皮肿腿蜂(S. brevicomis Kieffer)、柄腹茧蜂 (Spathius erythrocephalus Wesmael)、广肩小蜂(Eurytoma melanoneura Walker) 和 E. morio Boheman、丽旋小蜂(Calosota vernalis Curtis)和 C.agrili Nikol’skaya、旋小蜂 (Eupelmus aloysii Russo)、短颊金小蜂 (Cleonymus brevis Boucek)、长体金小蜂(Trigonoderus princeps Westwood)。国内外研究较多且生物防治效果较好的寄生性天敌为花绒寄甲(D. helophoroides)、川硬皮肿腿蜂(S. sichuanensis)、欧洲硬皮肿腿蜂(S. brevicornis)以及天牛卵长尾啮小蜂(A. fukutai.)[63]。在柑橘果园释放花绒寄甲成虫,防控效果可高达80%[64],川硬皮肿腿蜂对柳树上星天牛防控率可达43.63%[65]。
3 防治策略
3.1 严格林木检疫
天牛的自身扩散能力有限,远距离传播主要依靠苗木、包装材料及运输工具等人为活动,因此加强检疫是从源头上控制天牛传播扩散的有效途径。一是要根据虫害分布危害情况,确定疫区和保护区,严格禁止从疫区调运林木,阻断害虫传播途径。二是加强疫区林木调运的溯源管理,完善木材调运全过程检疫监管体系,实现疫区林木调运全过程可追溯、可监督。三是加强检疫除害方法研究,改进药剂熏蒸、药剂喷洒等传统除害处理方式,利用温度、低氧胁迫机制,推广智能热处理检疫除害监控技术,建立简单易操作有效的检疫除害新方法。四是加强国际协作,统一检疫标准,共同应对两种天牛的跨境传播风险,保障进出口贸易。
3.2 及时清理虫害木
光肩星天牛和星天牛耐性很强,能够在温度和氧气浓度很低的条件下生存,且主要藏于树干木质部内隧道越冬,因此在天牛危害严重发生区,需及时移除和处理虫害木,修剪受害枝干;对危害严重且无利用价值的被害树木进行彻底清除与无害化处理,以控制虫源扩散。但注意成虫期应避免砍伐天牛虫害树木或对虫害树木修枝平头,以防止人为扩散。
3.3 选用抗性树种
光肩星天牛和星天牛的寄主范围虽然广泛,但是对不同寄主有选择性差异。可结合本地生态和气候条件,选择既抗虫又适生的树种作为推广品种,比如杨树中冀秦1号杨和冀秦2号杨、豫林K-38号无性系、健杨、沙兰杨、欧美杨等杨树具有较强的抗虫性,可有效降低种群数量和危害程度。在造林规划中可按照一定比例种植非寄主树种、抗性树种和易感树种(诱杀树种),营造混交林,避免林带、林网、林地、林种单一化。或者按照一定的隔离带宽度设置高抗隔离带,比如在外围种植臭椿、刺槐、国槐、白花泡桐、毛泡桐、银杏、梧桐等高抗品种,形成生态屏障。
3.4 建立天敌栖息繁殖区
保护和充分发挥啄木鸟、花绒寄甲、寄生蜂等天敌的防虫治虫作用。通过挂设鸟巢招引天敌鸟类,促进天敌鸟类繁殖;在天牛严重危害区,通过释放肿腿蜂、花绒坚甲等天敌昆虫进行生物防治。因大部分昆虫需要取食花蜜和花粉才能产卵、繁衍后代,因此可在天牛发生区,适当种植一些有利于天敌生存的植物如十字花科、伞形花科等蜜源植物,这些植物可诱集大量的天敌栖息、生存和繁殖,可有效地增加天敌昆虫数量,提高控制害虫的能力。
3.5 加强监测预警
为更好防控天牛危害,前期监测十分必要。可在疫区或者威胁区固定监测点,开展预测预报,为防治提供科学的决策依据。整合现代技术和传统手段,在人工调查的基础上,建议结合卫星遥感、无人机航拍和地面监测系统,构建多维度一体化的监测体系,提升天牛的监测精度。同时结合历史发生数据,建立统一的虫情数据库或信息管理平台,实现监测数据实时共享和动态预警,提升对天牛虫情“早发现、早预警、早处置”能力。
4 结语和展望
在国内,光肩星天牛被列入危险性林业有害生物名单,在国际上两种天牛已成为全球重要检疫性害虫。因此,加强防控,不仅是应对国际上检疫的要求,也是保障国内林木健康的需要。综合现有的文献记录,这两种天牛的分布数据均为20年之前的数据,在全球气候变化和国际贸易发展浪潮下,两种天牛在中国的分布和危害情况是如何变化的,有待进一步深入调查和研究。
天牛类蛀干害虫生活隐蔽,防治难度大,“早发现、早反应”是防控成功的关键,传统人工调查方式成本高、有滞后性,在未来研究中应聚焦于声学传感器、遥感、AI图像识别等开发新型早期监测技术。同时利用大数据、气候模型和机器学习,构建精准的疫情扩散风险预测模型,评估气候变化(如变暖、干旱)对天牛种群动态和分布范围的影响,实现疫情精细化风险管理。
参考文献
[1] Keena M A, Sánchez V. Reproductive behaviors of Anoplophora glabripennis (Coleoptera: Cerambycidae) in the laboratory [J]. Journal of Economic Entomology, 2018, 111(2): 620-628.
[2]涂业苟, 金明霞, 王文辉, 等. 星天牛对江西薄壳山核桃的为害及生物学特性观察[J]. 中国植保导刊, 2024, 44(10): 59-63.
[3]佳昊, 黄波, 王德辉, 等. 光肩星天牛与星天牛种间竞争行为研究[J]. 环境昆虫学报, 2022, 44(3): 651-657.
[4]李呈澄, 任利利, 王立祥, 等. 生态自控光肩星天牛灾害的研究基础与展望[J]. 应用昆虫学报, 2025, 62(1): 31-41.
[5]周琳, 马志卿, 冯岗, 等. 天牛性信息素、引诱植物和植物性引诱剂的研究与应用[J]. 昆虫知识, 2006(4): 433 -438.
[6]安榆林. 光肩星天牛分子遗传多样性及PCR鉴定技术的研究[D]. 南京: 南京林业大学, 2004.
[7] R A Haack, F Hérard, J Sun, et al. Managing Invasive Populations of Asian Longhorned Beetle and Citrus Longhorned Beetle: A Worldwide Perspective[J]. Annual Review of Entomology, 2010, 55: 521-546.
[8] Wang L X, Li C C, Luo Y Q, et al. Current and future control of the wood-boringpest Anoplophora glabripennis[J]. Insect Science, 2023, 30(6): 1534-1551.
[9]刘德庆, 张天媛, 张晓丽, 等. 华北地区光肩星天牛空间分异性及驱动因素研究[J]. 农业机械学报, 2022, 53(1): 216-223+369.
[10] Yasser Alramadan, Mehmet Mamay, Shahid Farooq. Increased spread risk of citrus long-horned beetle [Anoplophora chinensis (Coleoptera: Cerambycidae)] under climate change in Türkiye: Implications for management[J]. 2025,190:https://doi.org/10.1016/j.cropro.2024.107090.
[11] KAPPEL A P, T R OTTER R T, KEENA M A, et al. Mapping of the Asian longhorned beetle’s time to maturity and risk to invasion at contiguous United States extent[J]. Biological Invasions,2017, 19(7): 1999-2013.
[12]刘月方, 魏国玲, 魏国英, 等. 杨树林光肩星天牛数量分布影响因子分析[J]. 内蒙古林业科技, 2012, 38(4): 60-61+64.
[13]温俊宝, 李友常, 夏乃斌, 等. 杨树光肩星天牛成虫扩散格局的研究[J]. 生态学报, 1998, 18(3): 47-55.
[14]李国宏, 高瑞桐, Michael T.Smith, 等. 光肩星天牛种群扩散规律的研究[J]. 林业科学研究. 2010, 23(5): 578-684.
[15] HULL-SANDE R S H, PEPPER E, DAVIS K, et al. Description of an establishment event by the invasive Asian longhorned beetle ( Anoplophora glabripennis) in a suburban landscape in the northeastern United States[J]. PLoS One, 2017, 12(7): e0181655. https://doi.org/10.1371/journal.pone.0181655.
[16]郭琳. 光肩星天牛不同越冬时期幼虫相关耐寒基因的表达规律[D]. 北京: 北京林业大学, 2019.
[17]韦美泽. 中国北方光肩星天牛的低氧适应性研究[D]. 北京: 北京林业大学, 2020.
[18] EPPO (2025) Anoplophora chinensis[DB/OL]. https://gd.eppo.int.2020-10-21/2025-11-14.
[19]苏筱雨, 李会平, 阎浚杰, 等. 桑天牛和光肩星天牛生态地理分布及无公害控制技术研究[J]. 林业与生态科学, 2020, 35(4): 393-398.
[20]王志刚. 中国光肩星天牛发生动态及治理对策研究[D]. 哈尔滨: 东北林业大学, 2004.
[21]李浩培, 吕飞, 毕拥国, 等. 中国林业重要蛀干害虫光肩星天牛研究进展[J]. 林业与生态科学, 2020, 35(1): 1-9.
[22]魏建荣, 赵文霞, 张永安. 星天牛研究进展[J]. 植物检疫, 2011, 25(5): 81-85.
[23]张树怀, 夏秀清, 苏红田, 等. 光肩星天牛发育起点温度和有效积温的研究[J]. 内蒙古农牧学院学报, 1995, 16(2): 45-49.
[24]杨志敏, 王西南, 姚文生, 等. 光肩星天牛世代分化及有效积温的研究[J]. 森林病虫通讯, 2000, 19(6): 12-14.
[25] Keena, M.A. Effects of temperature on Anoplophora glabripennis (Coleoptera: Cerambycidae) adult survival, reproduction, and egghatch[J]. Environmental Entomology, 2006, 35: 912-921.
[26] KEENA M A, MOORE P M. Effects of temperature on Anoplophora glabripennis (Coleoptera: Cerambycidae) larvae and pupae[J]. Environmental Entomology, 2010, 39(4): 1323-1335.
[27]陈辉, 周嘉熹, 邵崇斌, 等. 西北地区黄斑星天牛发生区域分析预测[J]. 西北林学院学报,1995, 10(2): 36-39.
[28]刘昭阳, 李玉蓉, 潘忠玉, 等. 光肩星天牛两型种群表型多样性分析[J]. 应用昆虫学报. 2016, 53(5): 1045-1057.
[29]周嘉熹, 杨雪彦, 王伟平. 黄斑星天牛预测预报技术[J]. 西北林学院学报, 1992, 3(7): 1-6.
[30]仲丽, 仇兰芬, 夏菲,等. 环境条件对光肩星天牛卵孵化的影响[J]. 安徽农学通报. 2019, 25(16): 83-84, 109.
[31] Melody A. Keena, Paul M. Moore, Gregg Bradford. Effects of Temperature on Anoplophora chinensis (coleoptera: Cerambycidae) Adult Survival, Reproduction and Egg Hatch[J]. Forests, 2021, 12(4): 432-432.
[32] Adachi I. Development and life cycle of Anoplophora malasiaca (Thomson) (Coleoptera: Cerambycidae) on citrus trees under fluctuating and constant temperature regimes[J]. Applied Entomology and Zoology, 1994, 29(4): 485-497.
[33]黄金水, 何学友, 丁泌, 等. 星天牛蛹的发育及成虫补充营养特性的研究[J]. 林业科学, 2003, 39(专刊): 148-152.
[34] CABI (2020). Anoplophora chinensis (black and white citrus longhorn)[DB/OL]. https://www.cabi.org/isc/datasheet/5556.2020-11-10/2025-08-21.
[35]李文杰, 邬承先. 杨树天牛综合管理[M]. 北京:中国林业出版社, 1992: 29.
[36]阎浚杰. “三北”地区光肩星天牛发生动态的研究[J]. 陕西林业科技, 1992(2): 62-66.
[37]高瑞桐, 李国宏. 中国光肩星天牛研究回顾及发展趋势[J]. 昆虫知识, 2001, 38(4): 252-258.
[38]杨忠岐, 王小艺, 张翌楠, 等. 以生物防治为主的综合控制中国重大林木病虫害研究进展[J]. 中国生物防治学报, 2018, 34(2): 163-183.
[39]王志刚, 阎浚杰, 刘玉军, 等. 西藏南部光肩星天牛发生情况调查报告[J]. 哈尔滨: 东北林业大学学报, 2003(4): 70-71.
[40] Quan-Cheng Zhang, Jun-Gang Wang, Yong-Hui Lei. Predicting Distribution of the Asian Longhorned Beetle, Anoplophora glabripennis (Coleoptera: Cerambycidae) and Its Natural Enemies in China[J]. Insects. 2022, 13, 687. https://doi.org/10.3390/insects13080687.
[41] Yasser Alramadan, Mehmet Mamay, Shahid Farooq. Increased spread risk of citrus long-horned beetle [Anoplophora chinensis (Coleoptera: Cerambycidae)] under climate change in Türkiye: Implications for management[J]. Crop Protection, 2025, 190: 107090. https://doi.org/10.1016/j.cropro.2024.107090.
[42] Xuezhen Ge, Shixiang Zong, Shanyong He, et al. Areas of China predicted to have a suitable climate for Anoplophora chinensis under a climate-warming scenario[J]. Entomologia Experimentalis Et Applicata, 2014, 153(3): 256-265.
[43] Byeon D, Kim S, Jung J, et al. Climate-based ensemble modelling to evaluate the global distribution of Anoplophora glabripennis (Motschulsky)[J]. Agricultural and Forest Entomology 2021, 23, 569-583.
[44] Zhou YT, Ge XZ, Zou Y, et al.. Prediction of the potential global distribution of the Asian longhorned beetle Anoplophora glabripennis (Coleoptera: Cerambycidae) under climate change[J]. Agricultural and Forest Entomology, 2021, 23: 557-568.
[45] Lyu Fei, Hai Xiaoxia, Wang Zhigang. A Review of the Host Plant Location and Recognition Mechanisms of Asian Longhorn Beetle[J]. Insects 2023, 14(3), 292. https://doi.org/10.3390/insects14030292.
[46]黄咏槐, 黄华毅, 钱明惠, 等. 星天牛寄主选择研究[J]. 环境昆虫学报, 2019, 41(2): 323- 328.
[47] FLø D, Krokene P, Økland B. Invasion potenial of Agrilus planipennis and other Agrilus beetles in Europe: import pathways of deciduous wood chips and Maxas[J]. EPP Bulletin, 2015, 45(2): 259-268.
[48] Henrik Sjöman, Johan Östberg, Johan Nilsson. Review of Host Trees for the Wood-Boring Pests Anoplophora glabripennis and Anoplophora chinensis: An Urban Forest Perspective[J]. Arboriculture & Urban Forestry, 2014, 40(3):143-164.
[49] Shim J Y, Jung J M, Byeon D H, et al. Evaluation of the spatial distribution of Anoplophora glabripennis (Coleoplorhora:Cerambycidae) in South Korea combining climate and host pant distruibution[J]. Journal of Asia-Pacific Entomology, 2020, 23(3): 646-652.
[50] Dang Y Q, Zhang Y L, Wang X Y, et al. Restrospective and analysis of an invasive wood-boring insect using native range data: the importance of host plants[J]. Journal of Pest Science, 2021, 94(3): 981-990.
[51] Lee S, Lee Y, Lee S. Population genetic structure of Anoplophora glabripennis in South Korea: invasive populations in the native range[J]. Journal of Pest Science, 2020, 93(4): 1181-1196.
[52]张宇凡, 党英侨, 王小艺. 基于气候和寄主因素的栗山天牛中国成灾和扩散风险评估[J].林业科学, 2022, 58(6): 96-108.
[53]嵇宝中. 中国天牛虫灾的成因、现状与对策[J]. 世界林业研究, 2001, 14 (4) : 50-56.
[54]罗立平, 王小艺, 杨忠岐, 等. 光肩星天牛生物防治研究进展[J]. 生物灾害科学, 2018, 41(4): 247-255.
[55]万涛, 矫振彪, 温俊宝, 等. 冬季大斑啄木鸟对光肩星天牛的选择性捕食[J]. 动物学报. 2008(3): 555-560.
[56]唐桦, 刘益宁, 马国骅. 宁夏地区光肩星天牛天敌种类调查初报[J]. 森林病虫通讯, 1996, 15(1): 30-31.
[57]任哲. 光肩星天牛柄腹茧蜂的繁育生物学特性研究[D]. 保定: 河北农业大学, 2024.
[58] Duan J J, Aparicio E, Tatman D, et al. Potential new associations of north american parasitoids with the invasive Asian longhorned beetle (Coleoptera: Cerambycidae) for biological control[J]. Journal of Economic Entomology, 2015, 109(2): 699-704.
[59]李飞. 光肩星天牛自然种群关键控制因子研究[D]. 沈阳: 沈阳农业大学, 2017.
[60]姜嫄, 张翌楠, 李志强, 等. 花绒寄甲防治旱柳光肩星天牛研究[J]. 北京农业职业学院学报, 2019, 33(6): 12-19.
[61]温玉洁, 赵文龙, 韩亮, 等. 花绒寄甲生物防治光肩星天牛研究[J]. 北京农业职业学院学报, 2022, 36(6): 65-71.
[62]张宇凡, 王小艺.星天牛生物防治研究进展[J]. 中国生物防治学报, 2019, 35(1): 134-145.
[63]黄咏槐,钱明惠,黄华毅,黄焕华. 星天牛防治技术研究进展[J]. 林业与环境科学, 2018, 34(4): 162-167.
[64]龚碧涯, 廖炜, 龙显耀, 等. 花绒寄甲对柑橘天牛的防控效果[J]. 中国植保导刊. 2023, 43(8): 61-65.
[65]杜开书, 周祖基, 杨伟. 川硬皮肿腿蜂防治柳树星天牛试验初报[J]. 安徽农业科学, 2006, 34(13): 3104-3105.
(a)光肩星天牛; (b)星天牛; (c)2种天牛鞘翅基部比较
图1 光肩星天牛和星天牛的成虫示意图[17-18]
Fig.1 The schematic diagram of adults of Anoplophora glabripennis and Anoplophora chinensis[17-18]
表1 光肩星天牛和星天牛发育温度阈值
Table 1 Developmental temperature thresholds of Anoplophora glabripennis and Anoplophora chinensis
天牛种类 | 最低发育阈值温度 (℃) | 全世代有效积温 (日度) | 年均温 (℃) | 参考文献 | ||
卵 | 蛹 | 幼虫 | ||||
光肩星天牛 | 9.7 | 10 | 1~5龄: 10; 高龄: 12 | 1264.2 | 3.9 | [24-26] |
星天牛 | 13 | 10 | 1~6龄: 10; 高龄: 12 | — | 10 | [31, 34] |
